

The XIX century will be called the century of Darwin (L. Boltzmann)
While most people have heard of Einstein and Newton and Feynman, Boltzmann is not a household name when thinking about famous physicists. Ludwig Boltzmann was a theoretical physicist extraordinaire who at the end of the XIX century, in that Vienna that was going to give so much to the world in the ensuing years, taught us a most interesting way of thinking in material terms about the structure of matter and abstract concepts like heat and energy. Spurred by his philosophical inclinations, in his latter years he wanted to transcend what he had done and thought, by looking at Evolution from the physical perspective. In this process he clearly absorbed much of Darwin at a time that darwinism was not as popular as it would become later: “… If you would ask me about my heartfelt conviction, whether the nineteenth century will be called one day the iron century or the century of the steam engine or the century of the electricity, I answered without any doubt it will be called the century of the mechanistic conception of nature, the century of Darwin…â€. There is little doubt from this statement that Boltzmann understood Darwin but there is also an inkling, if you know something about the work of each of these individuals, that he might have had a deeper insight than he let us know in his writings.
Physics and Biology share one challenge: the mechanistic understanding of the relationship between events that happen at the limit of our visual detection –the microscopic world- and what we can observe and sense i.e. measure (any act of perception is a more or less conscious measurement) at the macroscopic level. The way we do this is nicely put in a statement attributed to the physicist Jean Perrin, which suggests that one of the cornerstones of Science is the craft of revealing the invisible through the visible. In some respects this is what we do in Biology when we draw those diagrams that are meant to represent events supposed to happen inside cells. While some of them are probably accurate (and for accuracy on the basis of our current understanding of our molecular structural knowledge, see D. Goodsell visions of the cell: http://mgl.scripps.edu/people/goodsell/) others do not capture, yet, what they want to represent. And so, there is a two way road from the macroscopic to the microscopic. A topic of many talks in Biology is, we are told, that what we want to know is the relationship between the genotype and the phenotype, between the genes and the cell. However, behind this statement there is the dream of some sort of a linear relationship between both which has not and will not be found because 1) it does not exist and 2) this might not be the right question to ask. If you are an evolutionary biologist you spend a great deal of time relating genes to the structure of populations and therefore you know about the problems of simple linear models and of the slippery nature of quantifiable variables which are sometimes needed to deal with biological systems. However, it is precisely in the challenge of relating genes to, for the sake of argument let us say phenotypes, that the connection between Boltzmann and Darwin emerges and might provide some inspiration for today’s challenges.
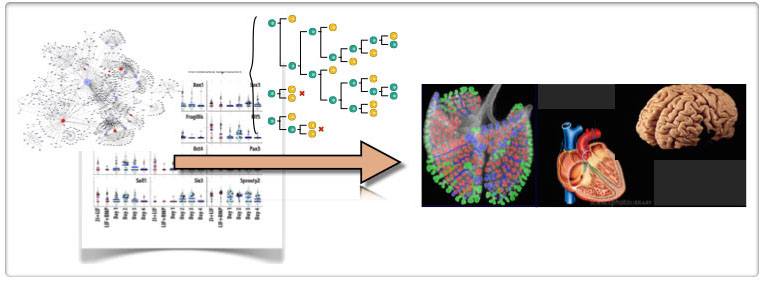
Figure 1. One of the big challenges in Biology is how to relate the events that are described by molecular networks with the organs and tissues that characterize the make up of an organism. Â It is obvious that cells and their lineages are the vehicles for this transformation.
The breakthrough of Boltzmann stemmed from his belief in the reality of atoms and their fundamental role in the understanding of physical systems. A belief it had to be since at that time it was impossible to penetrate the structure of a cell, let alone that of a molecule or an atom. Taking this view as a starting point, he developed a theory which provided a mechanistic explanation (watch it, not in the sense of the modern biologists i.e. figure 7 of your NSC paper, but rather, to quote my colleague Ben Simons, as a causal explanation for an observation) for observables like Pressure, Temperature or Energy. He showed how if one accepted the existence of atoms, one could derive these properties from the spatially constrained interactions between them. Since the number of molecules in a macroscopic observable is enormous (remember Avogradro’s number is 6.02 X10 23 molecules in a mole), even those who were interested in the subject, found it very difficult to comprehend how could one devise a mechanistic and mechanical way to connect these large numbers to the observables. If you were a committed newtonian you would have to calculate the trajectories and energies of every atom and its interactions with all the other atoms and then find a way to compute the total sum (or product) of the resulting numbers! The way forward, as Boltzmann saw, was assuming the reality of the atomic structure of matter, to perform a proper statistical analysis of the behaviour of ensembles of molecules in different conditions. He reckoned that with such large numbers, the connection between the elements and the properties of the system was through statistics –in its infancy at the time- and that under the simple conditions of an ideal gas, a statistical treatment of the kinetic relationships between individuals in populations of molecules (microscopic) would yield the macroscopic measurable (Pressure, Temperature, Kinetic Energy…); a proper treatment of the problem shows how the observables result from the constrained averaging of the individual variables. It was a deep insight that what mattered were the statistical properties of the population rather than the details of the individual behaviours which became averaged at the higher level. This work provided a solid foundation for the work of the Scottish physicist JC Maxwell who had calculated the distributions of velocities of an ideal gas on similar terms, thus laying a significant foundation for the kinetic theory of gases -this is why today we talk about the distributions of velocities and energies in physical systems as the Maxwell-Boltzmann distribution. But Boltzmann took the basic ideas of a statistical analysis of the structure of matter further and provided a material basis for that most elusive notion: Entropy (which in thermodynamic terms can be defined as the amount of energy, thermal energy, which is not available to do mechanical work). With apologies to the physicists (if any reads this) for the simplification, he envisioned matter as a problem in combinatorials of its constituents: a particular structure being one, and only one, of a huge number of configurations of its constituent elements. If that structure disappears, or changes, it means the system has acquired a new configuration and will search for the original one in the large space of all the other configurations. Not surprisingly it will find many ‘disorderered’ ones before finding the original one. Entropy, Boltzmann saw, is a measure of that number of non-structured configurations. He extrapolates this to the Universe and suggests Life as the chance result of a fluctuation in a small space of a large heat bath. It is these thoughts about the Evolution of physical systems that probably led him to consider darwininan concepts: “… The struggle for existence of the living beings is not a fight for basic materials—these materials are available in air, water and soil in sufficient quantities for all organisms—it is also not a fight for energy that is available in the form of inconvertible heat in every body but it is a fight for [negative] entropy, which becomes available by the transition of energy from the hot Sun to the cold Earth. In order to exploit this transition as much as possible, the plants spread out the incredibly large surface of the leaves and force the energy of the Sun before it falls down to the temperature of the Earth in a not yet understood way to perform synthetic chemical reactions that are still completely unknown in our laboratories. ..â€. Much food for thought here and I shall leave it for another time. Suffice to say that the deep gauntlet that lies in here was taken later by E Schrodinger who in his famous book “What is Life†discussed at length some of these notions and introduced the eye catching but misleading notion of negative entropy, free energy really (Gibbs or Helmholtz); he might have been influenced by his youth in Vienna studying Physics under the aura or the great Boltzmann.
 Â
Â
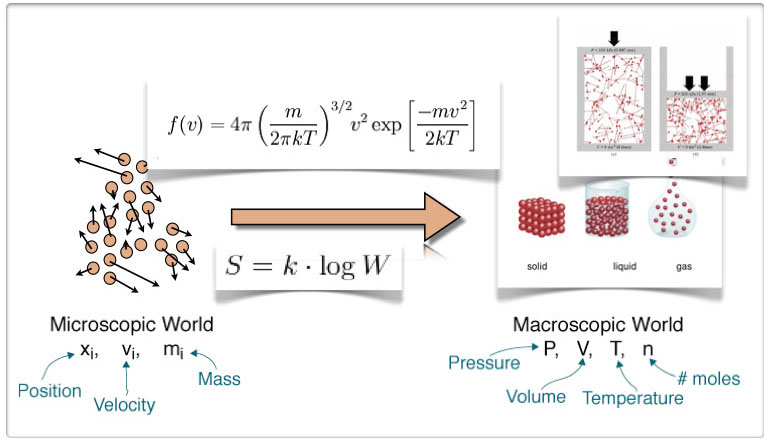
Figure 2. Boltzmann’s insights that allowed him to use statistics of the mechanical properties of the particles under several constrains to deduce the macroscopic properties of the system. In the process he provided a physical description of Entropy (S) in terms of the configurations of the system (W).
What does this have to do with where we are at the moment? What is the point of all this to modern Biology? The current challenge, as some of us perceive it, is not to see how genes generate a phenotype but to link the molecular and the cellular realms. To explain cellular activities (motility, change of fate, higher order structure and dynamics of cell populations, etc) in terms of their molecular underpinning. In all this and what has become a game changer is our ability to measure or, if you will, to see and then to measure, and to be able to do this at the level of individual cells. What we are getting out of this process is large amounts of data, information, that we are accumulating in databases that are more or less centralized and organized. What we are lacking is not just methods to process this information, but questions, conceptual frameworks to interpret what the analysis of the data (which is more data) yields. The question then can be reduced to how the myriads of genes, proteins and their interactions at one level, generate behaviours at a different scale. How do the macromolecular complexes that underpin cell movement  and shape, the structure of a tissue or the dynamics of a tissue in homeostasis, generate those observables?. In this work, there are two connected relations: from the molecules to the cell and then from the cell(s) to the tissue. This statement contains the implicit statement that THE CELL is a vehicle to link molecules to tissues and organs. The numbers of the game are very large (genes, transcripts, cells) and become larger if we consider single cells, which is becoming routine. It is here that the work of Boltzmann becomes an inspiration. The secret will be the averaging and the way biological systems do what physicists call coarse graining, will provide the understanding; but first we need to define the variables that need to be averaged and the calculations that need to be made. Progress is being made but it is slow because, unfortunately, the emphasis is still in mindless data collection and on the naïve belief that describing it is understanding.
It was probably this deep insight into the population averaging of the properties of very large number of components of a system that led Boltzmann to have an intuitive understanding of Darwin. After all, the importance of large numbers and their dynamics is implicit in Darwin’s theory of natural selection and becomes explicit in the postdarwininan interpretation as in the work is R. Fisher, S. Wright and others, genes play the role of the atoms, and statistics is not just central, but develops around these ideas. Qualities, phenotypes, arise from the multivariate statistics of the effects of multiple genes. It is interesting, as has been discussed by J Gunawardena that much Genetics was developed without an understanding of the molecular structure of the gene and that for many years, the gene was a mathematical entity  (Biology is more theoretical than Physics, Mol Biol Cell. 2013 Jun;24(12):1827-9).
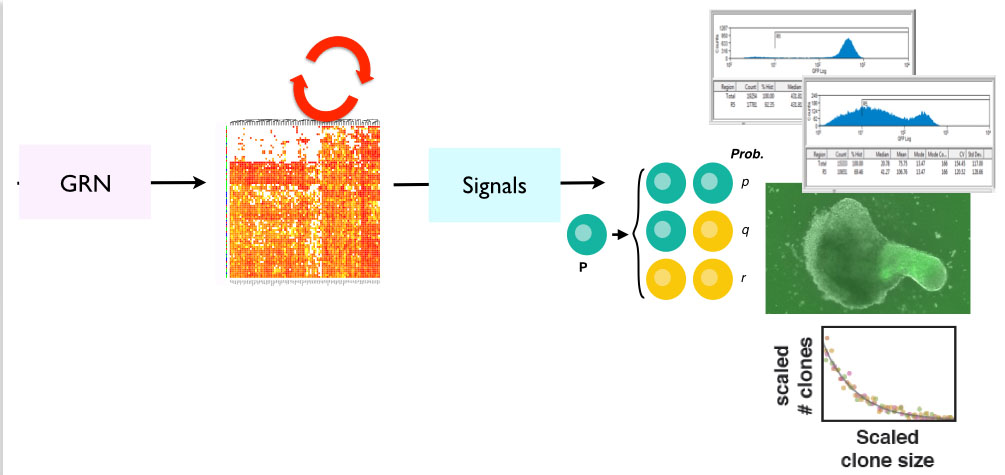
Figure 3. Outline for a statistical mechanics inspired solution to the problem (for further thoughts see references at the end). At the more microscopic level there are Gene Regulatory Networks (GRN) which generate dynamic (revolving arrows) patterns of activity at the level of single cells (intrinsic component). An interaction between these patterns and external signals (extrinsic components) generate patterns of fates at the cellular level that result in distributions of cell populations which are the result of distributions of gene expression in those populations. In turn these interactions across scales result in macroscopic structures. At the moment we do not know what these significant variables are nor what are their relationships but there are glimpses of this in the literature (see references at the end).
We need to look at Physics for inspiration and the current impasse needs, quickly, some new paradigms to move from description to understanding. The single cell analysis of developmental processes and, in particular stem cell populations has raised the possibility that statistical mechanics can offer a useful paradigm. What you have read for the last few minutes is a statement in support of such programme. But what we shall need is to define the macroscopic and the microscopic variables in a precise and meaningful manner. Then, progress will follow. Perhaps Boltzmann was right and the XIX century belongs to Darwin, as much as the XX belongs, at least in Biology, to the gene. In this series, the XXI should be the century of the cell and I hope that it does not take us 100 years to realize that to name and count genes and proteins is as futile a task as that which Boltzmann circumvented: to calculate the position and momentum of every particle of a gas. In many ways Biology is the unwritten chapter of statistical mechanics, the chapter that beckons at the end of any text book in the matter.
Darwin gave Biology a way to use the information that has been collated by naturalists in their collecting frenzy (which Darwin practiced in his early days). Today, instead of beetles and plants, we collect sequences and genomic landscapes and this is important and useful. However, the wonder of these objects and the useful information they contain should not deter our attention from the real task in hand which is to formulate the questions that will allow us to link genes (and epigenes) to cells and cell populations and through these to tissues and organs.
A brief list of related references (to build a field: the statistical mechanics of biological processes)
Karsenti E. Self-organization in cell biology: a brief history. Nat Rev Mol Cell Biol. 2008 Mar;9(3):255-62. doi: 10.1038/nrm2357 (E. Karsenti is a pioneer of the attempts to understand biological systems bridging the microscopic and macroscopic realms. He has done most of his work trying to understand how molecular ensembles generate cells which is a first step towards higher levels of understanding. His work is very influenced by I Prigogine).
Lander AD. Making sense in biology: an appreciation of Julian Lewis. BMC Biol. 2014 Aug 2;12(1):57. With Julian Lewis in mind, an insightful meditation of models in Biology.
Gunawardena J. Beware the tail that wags the dog: informal and formal models in biology. Mol Biol Cell. 2014 Nov 5;25(22):3441-4. doi: 10.1091/mbc.E14-02-0717. Models? What kind of models?
The next three references deal with the all important issue of time which is not dealt with here but is very important in linking molecular, cell and developmental biology:
Kicheva A, Cohen M, Briscoe J. Developmental pattern formation: insights from physics and biology. Science. 2012 Oct 12;338(6104):210-2. doi: 10.1126/science.1225182.
Kutejova E, Briscoe J, Kicheva A. Temporal dynamics of patterning by morphogen gradients. Curr Opin Genet Dev. 2009 Aug;19(4):315-22. doi: 10.1016/j.gde.2009.05.004.
Nahmad M, Lander AD. Spatiotemporal mechanisms of morphogen gradient interpretation. Curr Opin Genet Dev. 2011 Dec;21(6):726-31. doi: 10.1016/j.gde.2011.10.002.
The next four references discuss in an explicit manner the need for an approach based in statistical mechanics to understand the dynamics of cell populations in development.
Chalancon G, Ravarani CN, Balaji S, Martinez-Arias A, Aravind L, Jothi R, Babu MM. Interplay between gene expression noise and regulatory network architecture. Trends Genet. 2012 May;28(5):221-32. doi: 10.1016/j.tig.2012.01.006.
Garcia-Ojalvo J, Martinez Arias A. Towards a statistical mechanics of cell fate decisions. Curr Opin Genet Dev. 2012 Dec;22(6):619-26. doi: 10.1016/j.gde.2012.10.004
MacArthur BD, Lemischka IR. Statistical mechanics of pluripotency. Cell. 2013 Aug 1;154(3):484-9. doi: 10.1016/j.cell.2013.07.024.
Trott J, Hayashi K, Surani A, Babu MM, Martinez-Arias A. Dissecting ensemble networks in ES cell populations reveals micro-heterogeneity underlying pluripotency. Mol Biosyst. 2012 Mar;8(3):744-52. doi: 10.1039/c1mb05398a.
On the dynamics of cell populations:
Klein AM, Simons BD. Universal patterns of stem cell fate in cycling adult tissues. Development. 2011 Aug;138(15):3103-11. doi: 10.1242/dev.060103. This is an important insight from physics on the dynamics of cell populations.
Excellent text, almost to the end. Then, you write:
“The secret will be the averaging”
“we need to define the variables that need to be averaged”
I think this is one of the pitfalls of molecular biology, why it was often not able to explain cellular behaviour from molecular functions and disease from its molecular causes. The keyword is in your text: “the Maxwell-Boltzmann DISTRIBUTION”. Biologists have to give up on the average – also called arithmetic mean – to move to more complex description of their data. The field of drug development and testing moved past that. They had to, because humans are all different. But what we should realise is that even inbred animal models work with distributions. Any parameter, any initial condition, any trajectory is in fact better described as a distribution. Trying to remove variation from molecular and cellular processes at all cost (it is a bit different in some areas of physiology and neuroscience), assuming that it is only “noise” has been the curse of the last 3 decades. I think it did not just cause failure to understand and discover, it also produced a generation of narrow-minded and rigid scientists (and let’s be clear, everything I write here applies to me. This is all wishful thinking). We should interpret phenotypes in terms of differences of distributions. Very different distributions can provide the same average, while having different consequences. If we cannot compare raw distributions, we should at least consider the first four moments (for unimodal distributions, another widespread unsubstantiated assumption):
– mean
– standard deviation (and not SEM that does not reflect variation at all)
– skewness
– kurtosis
They all tell us something about the system.
End of the rant. I will just assume that by “average” you meant “distribution” 🙂
Scientific paradigms presume that space and time exist as a kind of vessel that houses atoms and molecules that make up the physical world. But space and time are concepts derived from physical observations a posteriori. They cannot be raised to a priori status to explain their own creation. This externalizes our understanding of the physical world while excluding our subjective reality that perceives it through sensory input to our central nervous system that recreates a virtual reflection of what is objectively “out there”. In other words we each relate to a subjective virtual reflection of the objective world as seen from our own unique perspective. A purely objective physical paradigm cannot account for this miracle. Cell processes are highly recursive because the active sites of protein enzymes essential to catalyze cell chemistry are not chemically altered in the process and they must also catalyze their own chemical synthesis, phosphate ions being the energy currency of exchange. It is a closed energy process that orchestrates cell chemistry from behind the scenes. This clearly indicates that there are archetypal energy patterns that intimately link Cells, Organs and the Host creature as an integral whole. It is universal archetypal energy patterns that constitute species and that have taken hundreds of millions of years to evolve. All this was unknown to Darwin and Boltzmann. There is no credible series of random chemical accidents that can create a new species. For example if both fruit fly parents have an eyeless gene their offspring will have no eyes, and yet after a few generations flies with eyes are born. The archetypal energy pattern of the species transcends and directs the chemistry of it members, while allowing of transient aberrations. For an alternate paradigm of how gene expression works see the article http://www.cosmic-mindreach.com/Gene_Expression.html. There are related articles on the website that introduce a practical universal methodology that applies to physics, quantum relativity, biology, and the human nervous system, and that is fully consistent with the huge accumulation of factual data gathered by traditional approaches to science. Science is at a crossroads and needs a new paradigm that is not dependent on language-based theoretical belief systems.