
Note: This is the second part of the last post and is not its final form. It will be updated and cleaned up in the New Year but wanted to share these thoughts with those of you who cared to read them before the treadmill catches up with me in the New Year.
The vis essentialis of Wolff, the Entelechia of Driesch, the new physical laws promised to Delbruck by Bohr, all found echoes in the famous book “What is life†by E. Schroedinger. This book, that meant so much to a few who went on to change Biology, deals with two questions concerning the physical nature of Life. The first one, central at the time, is the structure of the hereditary material. The second, more abstract and less emphasized in discussions of the book, focuses on the need to explore the thermodynamic basis of Living systems and in doing so it raises, again, the possibility that there might be new laws of Physics lurking in biological systems:
“Living matter, while not eluding the ‘laws of physics’ as established up to date, is likely to involve ‘other laws of physics’ hitherto unknown……. from all we have learnt about the structure of living matter, we must be prepared to find it working in a manner that cannot be reduced to the ordinary laws of physics. And that not on the grounds that there is any ‘new force’ or what not, directing the behaviour of the single atoms within a living organism, but because the construction is different from anything we have yet tested in the physical laboratory.
We must therefore not be discouraged by the difficulty of interpreting life by the ordinary laws of physics. For that is just what is to be expected from the knowledge we have gained of the structure of living matter. We must be prepared to find a new type of physical law prevailing in itâ€
By the time the book was written, the inevitability of Genetics as the key to unlock the chemical underpinning of living systems was widely accepted, and the double helix, the genetic code, the unravelling of the biochemistry of metabolism and the principles of gene regulation that followed, soon became the vindication of this statement. But, as Crick said, this was, and still is, all chemistry. New physical principles elude any questioning; perhaps, in the end, there are none.
A few weeks ago I attended a meeting of The Company of Biologists in Surrey (UK), “From stem cells to human developmentâ€. I went with a mixture of scepticism and curiosity. After all, how could one study human development? By studying I mean, not just describing it and comparing the normal with the pathological but rather doing the kind of work that, through the use of model organisms, has brought so much insight into the molecular and cellular mechanisms underlying the development of embryos. If we are just beginning to grasp how genes govern the development of mice and fish thanks to experimental intervention, how are we going to do the same with humans? It is not just that the material for these studies is difficult to obtain, it is that, with all reason, we need to be mindful of the ethics of this work. Such thoughts were in my mind fuelling low expectations. The meeting turned out to be, for me, a great surprise and the answer to many of my questions, I should have known, laid hidden in the title of workshop through stem cells to developmentâ€.
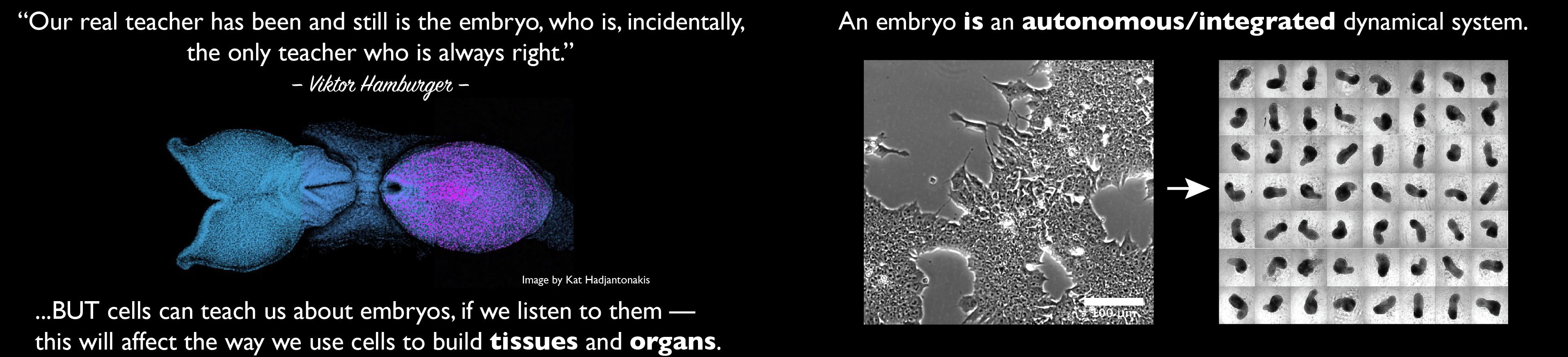
The meeting was a series of examples of what is becoming a clear fact to those in the know: cells are the vehicle between the genes and the organisms. Cells transform the instructions that lay dormant in the genome not just into proteins but into shapes and complex multicomponent forms. The structure of cells, and not just their physical organization but also their computational structure, drive their assembly in the macroscopic arrangements of different cells that we call tissues and organs, and this manifests itself, more than anywhere else, in the surprising self organizing activity of stem cells, embryonic and adult. Take cells with the right potential, place them in the appropriate culture conditions, ignite them with a signal and a genetically driven process will be unleashed that will transform a sequence of nucleotides into a multicellular structure. And the meeting showed us how human eyes, neocortexes, intestines, lungs and blood, emerged from stem cells, embryonic stem (ES) cells. Watching these unfold it is difficult not to think of these processes as manifestations of the vis essentialis and see in these cultures the opportunity to tackle its physical, or if you want to be more conservative, physical-chemical nature. And it is in watching these wondrous processes that the possibility of novel physical principles lurks again in the background.
The notion that cells derived from embryos have a self organizing activity had been known, but perhaps not appreciated, for some time. Thus, Holtfreter and Barth (discussed in a modern light by Hurtado and de Robertis 2007 Dev Biol. 307, 282-289) had observed that animal cap cells from Ambystoma maculatum salamanders, will differentiate autonomously into structures which resemble anterior cortex and develop eyes in culture. Furthermore, attempts to understand limb development and patterning contain numerous reports of mesenchymal cells, jumbled up and wrapped in ectodermal coats, generating digit like structures with recognizable identities. In the premolecular era, this type of experiment was the bread and butter of the experimental embryologist but though guiding much developmental biology, at the time there was little chance of understanding them. Over the last twenty years the application of the methods of classical genetics to development and pattern formation have yielded a catalogue of genes associated with particular processes. In this endeavour you remove a gene, look at the consequences for the organism and then try to work out what was the job of the gene in the process that has gone awry. Connections between genes are worked out through the process of epistasis. While this works very well with linear systems and particularly with metabolic routes, things can get out of hand with complex processes involving non linear systems like cellular machines, or processes like most of development and pattern formation. In particular, it is possible to find molecules that will induce nervous system from ectoderm or digits out of mesenchyme but, what about the process itself? But there is more to a process than its outcome; in fact the process is more interesting than the outcome. Is there a way to tackle its dynamics (the vis essentialis), the way cells proportion tissues and organs (entelechia)? Here perhaps, one needs to take a page (or a piece of the blackboard) of Richard Feynman who famously said: “that which I cannot build I do not understandâ€. This has been taken much at heart by synthetic biologists who ever since the ‘repressilator†(Elowitz, M. and Leibler, S. A synthetic oscillatory network of transcriptional regulators. Nature. 2000 Jan 20;403(6767):335-8) have been testing the information processing capacity of genetic circuits. But, in terms of Development, we are very far from being able to synthesize the very complex circuits and networks that drive the emergence of organs and tissues. Enter the cell.
Over the last five years there have been an increasing number of reports highlighting the ‘self organizing’ activities of cellular ensembles derived from stem cells. In work pioneered with intestinal stem cells, two cells from adult intestines can give rise in culture to structures that resemble very much in terms of composition and organization, the villi that configure the mammalian intestine. The same has now been shown to be true for other tissues, including lung and liver. However, in an extreme version of these experiments, the late Y Sasai (De Robertis EM. Yoshiki Sasai 1962-2014. Cell. 2014 Sep 11;158(6):1233-5) and his colleagues were able to, under specific culture conditions, coax ES cells to develop into retinas and neocortex. What is more, if the ES cells were from mouse, the emerging structures were sized as in a mouse but, if the starting point were human ES cells, the end point has the size of a human retina (Sasai Y, Eiraku M, Suga H. In vitro organogenesis in three dimensions: self-organising stem cells. Development. 2012 Nov;139(22):4111-21. doi: 10.1242/dev.079590). What else do you need to think about a driving force that is tailored to a particular species and which is able to asses in such a precise manner its final size and proportions? The meeting in Surrey had its share of these phenomena: human ES cells proliferating and turning into intestines, lungs and blood, which highlighted the cells as the architects of the organism through the interpretation of genetic programmes. But how do we get at the physical basis of these structures? Are there any new principles or physical laws behind these observations?
I do not think that self organization is the right notion for what is happening in these experiments; in some way what these cells do is, principally and certainly initially, to organize; however, as it is likely that everybody understands the term self organization, I shall stick to it – though in places I would rather remove the self: the ensemble self organizes, a cell organizes. In this context, there is something important, perhaps fundamental, we do not yet grasp –and need to come to terms with- in the fact that it has to be stem cells that do this and that their label of origin determines the final structure. The most straightforward interpretation would be that what stem cells can do and do remains deeply buried in their genetic programmes and that this is what fuels their organizing potential. Experiments in Drosophila shed some light on this. The Eyeless gene is at the top of a genetic hierarchy which controls the development of an eye. Eyeless is conserved in vertebrates, Pax6, in terms of structure and function -though Pax is involved in several functions, it is also involved in several aspects of eye development. Surprisingly, expression of Eyeless in any cell of the developing fruit fly will lead to a compound eye and, in this context, Pax6 will do the same thing. The simplest explanation for this is that Eyeless drives a deterministic programme for eye. Pax6 will do pretty much the same thing i.e. if expressed in Drosophila it will do the same as Eyeless. What this must mean is that Eyeless and Pax6, which are transcription factors, elicit a programme which in Drosophila’s software will produce a compound eye, the eye of Drosophila. There ought to be a homologous programme in vertebrates as Pax6 is required for the development of the eye. This is not self organization, but highlights that once a programme is initiated in a cell or a group of cells, it will be followed to term. It is maybe that some of what is going on in the cultures of stem cells have a component of this: a programme gets activated which in a deterministic manner will lead to the particular structure. The self organizing component comes into the picture the minute that there are different cells in the culture which now will not lose their way and will assemble themselves into specific structures. It is interesting and ill understood that this organization requires a 3D organization. However, it is important to realize that in this self organizing potential and much of what we can do is to steer this potential with alchemic precision. However as Jeremy Gunawardena once pointed out to me, there is hope since Chemistry is Alchemy with numbers!
Many questions emerge from these observations as well as many experimental possibilities but, new physical laws? New physical principles? Unlikely. Nonetheless and without getting too philosophical, there are two issues where Biology and Physics meet in these experiments which might lead to new notions or conceptual frameworks about the nature of space and time, what I would call the nature of biological space and time. Enough space left to just outline these and, paraphrasing Fermat’s margin, state that there is much to discuss on this, but not enough room here (or patience left in the reader) so, I will just make a few statements. The experiments with stem cells provide a system to determine how cells measure space i.e why mouse and human ES cells will each produce structures with sizes appropriate to their genetic blueprints? Where is this encoded? How is it decoded and executed? How do adult stem cells keep homeostasis of the size and shape of tissues? How do cells create such defined forms and shapes with a high degree of reproducibility? In an interesting observation, aggregates of ES cells have a critical mass to develop into specific patterns: above it, chaos; below it, inactivity. How do they know? How do they sense? But in addition to Space, there is also Time. And it is in this notion that new concepts, perhaps principles, will emerge. Time is central to biological systems as their dymamics, at any scale, is a most intrinsic property. Time, as is well known to physicists, is the most subjective of variables (as the physicist Sean Carroll puts it paraphrasing St Augustine : “I know what time is until you ask me for a definition about it, and then I can’t give it to you.”) however, biological time is not related in any simple manner to astronomical or sidereal time and when we do make this correlation we might be making a mistake. Take the process of somitogenesis (somites are the building blocks of vertebrates} which is run by a molecular oscillator whose elements are conserved across species but whose period is different in different embryos. But this difference is in astronomical time, perhaps from the perspective of the ‘system’ it does not matter, they are the same. How should we think about this? We can see these transformations in many processes which are run by the same molecular networks but take different astronomical times in different organisms. Is there a difference between the time of the networks and astronomical time? Time, as measured by the activity of genetic circuits (a network is not a circuit), is important for the correct decoding of ‘morphogen gradients†and is probably encoded and created by the activity of those circuits. If there is a process in which time is central it is gastrulation in birds and mammals. Furthermore, here, the convergence of space and time is the essence of the process. In these embryos, gastrulation is associated with a structure called the Primitive Streak: cells get progressively drawn into a groove through which they transit to generate the primordial of the different organs. There is an order: extraembryonic, blood, endoderm, heart, somatic…..what determines what a cell does is not its position in the embryo but the time and the order at which it enters the streak. There is a temporal programme which must be written in the gene networks and the circuits they produce. As JA Wheeler said, “time is the way to ensure that things do not happen all at onceâ€. Nowhere is this more clearly stated than in many biological processes and most and best of all in the process of gastrulation in birds and mammals. It is in understanding how genetic circuits generate and interpret time that many new insights will come about in biology and how we might find some novel physical notions (and its relationship to space). It is also likely that this is the vis essentialis and Entelechia.
There is a tinge of engineering in the way we are handling ES cells, but of XVIII and XIX century engineering in the sense that we are tinkering with something we do not quite understand and somehow getting it to work. However, unlike mechanical, civil or chemical engineering where humans run the system, in this engineering of cellular organization, the system rules and runs the scientists, using a blueprint that, for the time being remains hidden in the deep cellars of the cells –to emphasize that we really do not know where it lies.
Interesting and exciting times ahead as accessing that blueprint will reveal new principles and mechanisms, if not physical, certainly biological. Not just the notion of how cells generate time, their time, but also the averaging of fluctuations at any level at the higher level of organization e.g molecular to cellular, cellular to tissue. A new kind of engineering indeed.
Â
Thanks for one more excellent text, Alfonso. I was at the same meeting and shared the sense of marvel when seeing all those emerging cellular structures from human cells. The self-organisation capacity of tissues from “groups” of cells (or “ensembles”, a term that you also used – like orchestras) is certainly revealing an “hidden” layer of organisation, that has to be contained in the genetic information. As you refer in your Boltzmann post, we might soon be able to decipher this layer from the large datasets that are being generated.
But I have to add two points to the discussion:
1- I do not think that only “stem” cells can “self-organise”. I think you are using the “stem” word in the large sense that includes progenitors (proliferating), and this might be more adequate. But, remember that a “normal” capacity of any epithelial cell is to establish cellular bridges (junctions) with neighbouring epithelial cells and forms monolayers or “cysts”, a behaviour that is “self-organised”, and that does not necessarily involve “stem” cells. And this epithelial “property” is central to all “morphogenetic” movements in the embryo.
2. The point about gastrulation, in which you wrote that there is an order – what determines what a cell does is not the position but the time…”. Here, I have the feeling that we are all (wrongly) “eliminating from the equation” the fact that cells are fast dividing. We are not dealing with a single entity that is there “passively waiting and counting” time. No cell in the gastrulating embryo “lives” more than 10 hours (in the chick this was measured in Sanders, EJ, 1993- “2 hours at caudal levels of both the epiblast and mesoderm, to an upper value of approximately 10 hours in the rostral regions of the primitive streak…). So, the challenge is to understand how the genetic information is “prepared” to deal with these “jumps” in the lineage, and how new cells “know” at which point of the lineage they are. How are newcomers incorporated in the orchestra?
Finally, I like very much your effort to put time in the equation and your point that we need to understand how genetic circuits generate and interpret time. But I will not follow on the “speculation” that this is where we might find novel physical notions… cells count time using chemical reactions and these are “governed” by the known laws of physics, isn’t it? Or am I being too naive?