
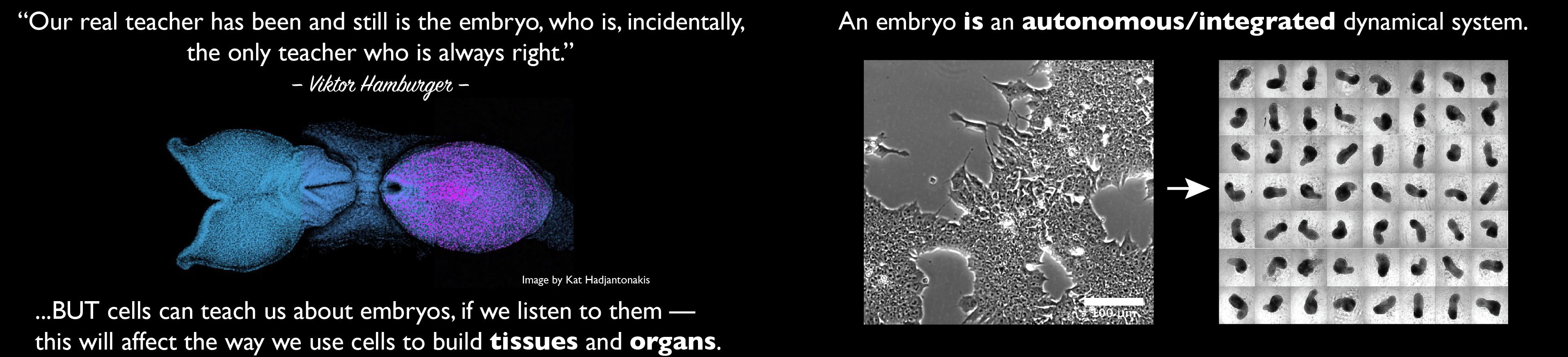
Over the last year a trend has emerged, highlighted by several articles, to defend model systems, specifically in Developmental Biology. The reason for this is a perceived (and real) threat in the funding of research in yeast, C. elegans, Drosophila, sea urchins and other systems e.g ascidians or arabidopsis (the issue of the animalcentric view of developmental biology is an interesting one) despite their research potential supported by many and obvious substantial contributions to our understanding of biological systems. One of the latest in this series of statement was published recently (1: https://dmm.biologists.org/content/10/12/1381). The authors do an excellent exposition of the contributions of several model systems to basic Biology and the impact that they have had on biomedical research, as a way to justify the need to continue the support of their research base. Surprisingly, the authors don’t include vertebrate models and Xenopus workers might query this omission. However, in addition to defend the aforementioned systems, they take issue with emerging research in Stem Cells, stating at the outset that “those who do not know the history of yeast, flies and other non-vertebrate organisms and their contributions to biomedicine may consider such studies as unworthy of participation or of funding. Given the growing excitement about human stem-cellbased cell culture systems, particularly organoids, some may argue that organisms that do not recapitulate all of the complexity of humans will cease to be useful in the near future. This line of thought is not only incorrect but could hamper scientific progress” (1). In a rather barbed comment towards the end of the essay, the authors state “We would go further to argue that organoids will not and should not replace non-vertebrate model organisms as discovery tools” (1) Here he authors refer directly to an article in the special issue of Development on organoids (2) in which it was argued exactly what is the theme of this post, that there is no way to separate the two fields and that putting them together promises much for the future.
As someone who is aware of that history and also works with Embryonic Stem Cells (ESCs) (DOI) I say that these statements are wrong and that to pitch research with Stem Cells and Organoids in contrast to that with model systems is not helpful. The issue is not about Stem Cells/Organoids versus model systems but about good and bad Science; and there is a lot of both in model systems and in Stem Cells, though I would say that from a strictly Science based perspective, right now, there is a lot of very good Science with model systems (DB) and a lot of bad science with Stem Cells at the moment. Issues of significance are a different matter. When making these arguments we should bear in mind that in comparison with model organisms, Stem Cells/Organoids are barely at the start of their historical life. In some ways Stem Cells, as an experimental system, are having the same disruptive effect that, say, Drosophila had when it erupted into a field dominated by marine invertebrates at the beginning of the XX century (3) and yet, history proofs that Drosophila, once it found its bearings, made very significant contributions and insights (3, 4). Difficult to compare historical periods but we should wait for time to make its mark in new fields of research. The discussion is not only about the quality of the Science that these areas of research can produce, it is also a matter of questions and systems – will expand below. Notwithstanding these considerations, I want ti insist that it is not helpful to create confrontations, particularly when research in Stem Cells has always been an integral part of Developmental Biology and therefore, to try to present it as something new and competitive is to deny an essential part of the field and systems that the authors try to defend: Developmental Biology. Our understanding of cell lineages in many organisms e.g leeches, and systems e.g the nervous system in all metazoan, of gametogenesis, homeostasis and regeneration is founded on the biology of Stem Cells. We should celebrate and welcome this. It is true that over the last ten years the field of Stem Cells has undergone a renaissance (as did Drosophila in the 1980s after a hiatus of almost 40 years) and that some people want to present it as something new, but it does not work. Research with Stem Cells is Developmental Biology and what has changed is that in their renaissance, Stem Cells and, in particular Embryonic Stem Cells (ESCs), have become a new model system for Developmental Biology.
Nobody will deny that the model systems highlighted by the authors have played a key role in modern Biology. In particular they have been instrumental in revealing the elements needed for biological systems to build themselves and to function. The genetic and molecular screens of the 1980s and 90s uncovered a set of agents that, although we did not know at the time, turned out to be elements of a universal tool kit for the construction of living systems, and many have been shown to be central targets of disease (5, 6). This work has led us to appreciate the critical role of transcription in cell fate assignments, that of cell biology in morphogenesis and how the combination of the two contributes to the building of tissues and organs. In learning about the universality of many of these elements we are uncovering that they form conserved modules whose functioning we still do not understand; continuing work on these systems is making progress to understand these modules. This is the basis for much of the arguments as those presented here (1) and it is right. However, Science, even today is an issue of questions and systems i.e. one should not work on a system simply because it is experimentally sound, because it yields results, because many experiments can be done or because it has a glorious past. One should work on a system because it is suited to ask, and maybe answer, specific questions and one should match question and experimental system. In this regard, invertebrates have many features in common with vertebrates but there are features of vertebrates that do not figure in the Biology of invertebrates. For example, the large scale cellular rearrangements that characterize most vertebrate embryos do not find a correlate in Drosophila or C. elegans, not to mention issues of homeostasis which only arise in the life spans of vertebrates. The issue of Dlp and Hippo that the authors raise in their essay (1) echoes the actions of hormones and chelons, but it is clear that the time and length scales characteristic of vertebrates require studying them in vertebrates. Interestingly, the issue of the regulation and function of Hippo –very much a flavor of the times- might not be the best example since it appears that Hippo can be linked to just about anything and everything and that one would always have to make such studies in a specific context.
At a most significant level, the fundamentally genetic work with model organisms, does not address the question of how cells build tissues and organs and this is because a gene-centric approach can’t do that; I hasten to add that the fact that there is a gene involved in a specific process and that we might be able to interpret gain and loss of function phenotypes does not count in my book. To understand how cells (genes is more difficult) build tissues and organs requires a very different approach from the one we have been using until now. You don’t understand how a car works simply by removing pieces and running it –sometimes it will crash, others it will not start or run slower- you learn how it works by building it and testing what you have built or are building the necessity and sufficiency of every piece (see the famed ‘Can a biologist fix a radio’ ref 7)-. The case of yeast and the cell cycle is an interesting one in this regard. Some of the most salient insights in this field have not come only from mutagenesis and epistasis but from a combination of modelling and engineering in which the order of the system is challenged and reorganized (8). This has led to discovering minimal systems, the functional meaning of redundancies and of modules with functional sense i.e going beyond genes. Behind these findings lurk principles and designs (see the interesting discussion presented in 9 and 10). Reengineering or engineering a system from the bottom up tells us much that Genetics cannot do. This kind of work is very difficult to do with model systems because they are not suited to this. In the case of some invertebrate models systems because they are too hardwired and one cannot create them from scratch. In the case of vertebrates because of their complexity challenges this type of approaches. Enter Stem Cells and, in particular ESCs as a new model system in Developmental Biology one in which we can ask how cells build tissues, organs, embryos.
 ESCs are a derivative from early mammalian embryos with a huge potential to enlighten our understanding of how cells build embryos, tissues and organs. It is true that it is early days, that there is a lot of poor work and a great deal of salesmanship of mediocre science at the moment, but this should not hide the fact of what they offer if they are used correctly, with high standards and a critical eye. And organoids, mentioned in (1) are an excellent example of what these cells (and also Adult Stem Cells (ASCs)) can do and how much we can learn from them about details of Biology that are not accessible in any other way. But, we need to use them judiciously and critically. Some of the arguments for this have been made before (https://amapress.gen.cam.ac.uk/?p=2024 and https://amapress.gen.cam.ac.uk/?p=996 ) and I shall not repeat them here, but will emphasize that Duronio et al are right that Stem Cells and organoids not a substitute for Model Organisms, they are a new experimental system with possibilities that are just beginning to be explored and whose full potential remains to be explored. They are not going to replace vertebrate model systems because the kind of questions that they ask are different and complementary but they sure will change the way we view vertebrate and, in particular mammalian development. Look at the work of a small group of people and their collaborators -J. Briscoe, S. Lowell, V. Wilson, J. Brickman, P. Zandstra, A. Brivanlou and E Siggia, A. Warmfklash, Fu, M. Lutolf, A. Grapin-Botton amongst others- that are exploring this avenue. There are also people who are using developmental biology to pioneer the development of regenerative medicine with ESCs, the work of J. Wells and J. Spence comes to mind. Here it is this dialogue between Developmental and Stem Cell Biology that matters. And in the area of adult organs, optic cups, minibrains and intestinal organoids have been leading a revolution in our understanding of precisely this question of how cells build organs and how we can harness this potential for biomedical research (11, 12). If one looks at this work, one will see that there is nothing to fear and much to be gained by promoting research with Stem Cells IN the context of Developmental Biology. To paraphrase a well known quotation: Stem Cells without model systems are lame, Model systems without Stem Cells are blind.
ESCs are a derivative from early mammalian embryos with a huge potential to enlighten our understanding of how cells build embryos, tissues and organs. It is true that it is early days, that there is a lot of poor work and a great deal of salesmanship of mediocre science at the moment, but this should not hide the fact of what they offer if they are used correctly, with high standards and a critical eye. And organoids, mentioned in (1) are an excellent example of what these cells (and also Adult Stem Cells (ASCs)) can do and how much we can learn from them about details of Biology that are not accessible in any other way. But, we need to use them judiciously and critically. Some of the arguments for this have been made before (https://amapress.gen.cam.ac.uk/?p=2024 and https://amapress.gen.cam.ac.uk/?p=996 ) and I shall not repeat them here, but will emphasize that Duronio et al are right that Stem Cells and organoids not a substitute for Model Organisms, they are a new experimental system with possibilities that are just beginning to be explored and whose full potential remains to be explored. They are not going to replace vertebrate model systems because the kind of questions that they ask are different and complementary but they sure will change the way we view vertebrate and, in particular mammalian development. Look at the work of a small group of people and their collaborators -J. Briscoe, S. Lowell, V. Wilson, J. Brickman, P. Zandstra, A. Brivanlou and E Siggia, A. Warmfklash, Fu, M. Lutolf, A. Grapin-Botton amongst others- that are exploring this avenue. There are also people who are using developmental biology to pioneer the development of regenerative medicine with ESCs, the work of J. Wells and J. Spence comes to mind. Here it is this dialogue between Developmental and Stem Cell Biology that matters. And in the area of adult organs, optic cups, minibrains and intestinal organoids have been leading a revolution in our understanding of precisely this question of how cells build organs and how we can harness this potential for biomedical research (11, 12). If one looks at this work, one will see that there is nothing to fear and much to be gained by promoting research with Stem Cells IN the context of Developmental Biology. To paraphrase a well known quotation: Stem Cells without model systems are lame, Model systems without Stem Cells are blind.
And as long as we don’t hype the issue and provide perspective, we should not shy away from the statement that understanding the make up and development of human beings is an important aim of Developmental biology. The reasons for this could be sheer curiosity or the thought of applications to disease or regenerative medicine. However, there is no ethically sound model system to study human development. The last few years have shown that while mice are a good proxy for much that is human, the only way to study humans is with humans: the work with minibrains and intestinal organoids, highlights this. And it is here that, hopefully on sound grounds, ESCs and ASCs will become a most important tool in this endeavour because what we are learning from the work with mouse cells is that if properly used, they teach is things. Not everything that happens in an embryo happens in the dish but all that happens in the dish can happen, and often happens in the embryo.
It seems to me that what Duronio et al grapple with is not really Stem Cells and Organoids but something more mundane but perhaps also more fundamental: the squeeze in funding, the large number of researchers and the average high quality of research that makes striving for that funding a challenging task. From this perspective, we should not lose sight that Stem Cells face the same problems than Drosophila. The reason is that funders often want to hear that one is going to cure a disease and not to find something fundamental; this applies to model systems as well as to Stem Cells. Now what is important is that we shall not convince funders by denying what is obvious (the possibilities opened by Stem Cells and Organoids) but by highlighting how Stem Cells and Developmental Biology are not only very related but are, as discussed above, complementary. We should also remember that the discoveries Duronio et al highlight were made in a very different environment than the one we worked on today when, perhaps (I believe certainly) it was easier to discern what was important from what was anecdotal or simply interesting. This is difficult today when every week we are confronted by ‘great’ discoveries which strive to be seen in a rain of technical prowess.
Stem Cells as a model system are here to stay and should be seen as a complement and not a competitor in our current arsenal to understand Nature. Is is interesting to reflect on the observation that one of the great achievements of Development journal has been to create a highly successful section on Stem Cells! What we need to do, though, is raise our game, find good, difficult questions and ways to address them. We should not be complacent and think of experiments simply because we can do them or of systems because we can use them. We should be bold and look for new avenues an systems. Also, we need to admit that much has changed and is changing in the environment in which we do Science and that what worked in the past, does not work today. We need to change and adapt. We need to accept that we work in a very different environment with very little room for the scientist/craftsperson of the XIX and XX century (https://amapress.gen.cam.ac.uk/?p=1537). What we need to do is to find ways to use much that is good in this environment and invent new ways of doing Science. In the context of the essay that triggers this post (1), we need to avoid justifying classical model organisms on what they have done in the past and find ways to make sure that they contribute to the future. Biological systems are such that there will always be a new gene, a new organelle a new relationship to be found, like the collections of the XVIII and XIX centuries were putting together animals and plants. What we need to make sure is that we find the ‘finches’ and have the eye to recognise what they tell us.
References
- Duronio, RJ., O’Farell, P., Sluder, G., Su, TT. (2017) Sophisticated lessons from simple organisms: appreciating the value of curiosity-driven research. Dis. Model Mech. 10, 1381-1389.
- Huch, M., Knoblich, J., Lutolf, M. and Martinez Arias, A. (2017) The hype and hpe of organoids. Development 144, 938-941
- Kohler,RE. (1994) Lords of the fly. The University of Chicago Press.
- EA Carlson “Mendel’s legacy’ Cold Spring Harbor 2004 .
- Anderson, KV and Ingham, P. The transformation of the model organism: a decade of developmental genetics. Nature Genetics 33, 285-293.
- Ugur, B., Chen, K. and Bellen, H. (2016) Drosohila tools and assays for the study of human disease. Dis, Model and Mech. 9, 235-244.
- Lazebnik, Y. (2002) Can a biologist fix a radio? Or what I learnt whie studying apoptosis. Cancer Cell 2, 179-182.
- Coudreuse, D. and Nurse, P. (2010) Driving the cell cycle with a minimal CDK control network Nature 468, 1074-1079
- Atay, O and Skotheim, J. (2014) Modularity and predictability in cell signalling and decision making. Mol. Cell Biol 25, 3445-3450
- Mellis, IA and Raj, A. (2015) Half dozen of one, six billion of the other: What can small- and large-scale molecular systems biology learn from one another? Genome Res. 25, 1466-1472.
- Sasai, Y. (2013) Next-generation regenerative medicine: organogenesis from stem cells in 3D culture. Cell Stem Cell 12, 520-530.
- Bredenoord, AL., Clevers, H. and Knoblich, J. (2017) Human tissues in a dish: the research and ethical implications of organoid technology. Science 355, DOI: 10.1126/science.aaf9414
I am a Teacher. your article is help me much in understanding concepts.
Thanks for this great article